

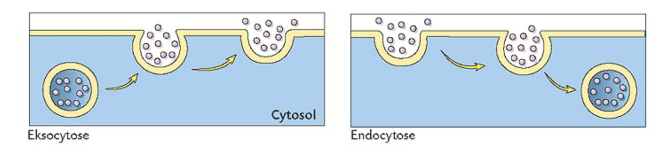
- Eksocytose
Intracellulære vesikler smelter sammen med cellemembranen og frigjør innholdet sitt til ekstracellulært rom
Eksocytose tilfører lipider og proteiner og gjør at cellemembranens areal øker. - Endocytose
Opptak av innhold fra ekstracellulært rom ved innvaginering av cellemembranen
Endocytose gjør at cellemembranens areal minker.
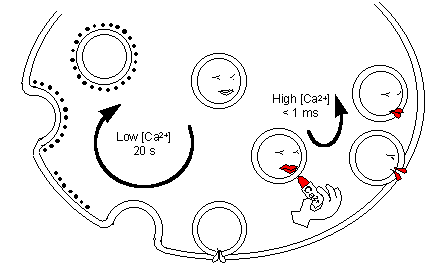
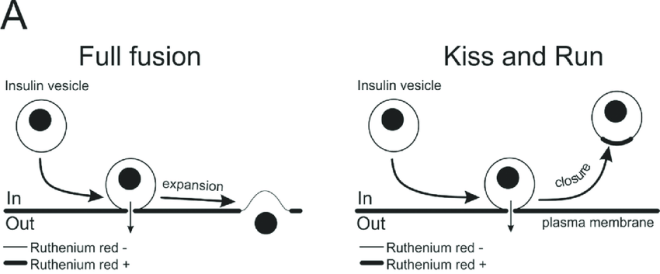
Kiss and run
Cellemembranens hvilespenning er på ~-70mv. Vi finner Na+ hovedsakelig utenfor og K+ innenfor. Det er flere K+ lekkekanaler enn for Na+. Når K+ lekker ut blir det mer negativt inni cellen enn utenfor (likevektspotensialet for K+ er ~-90mv). Hvilespenningen er ~-70mv på grunn av Na+ som lekker inn. Na+/K+-pumpen opprettholder hvilepotensialet.
- Intracellulært
K+ og andre organiske anioner - Ekstracellulært
Na+, Cl-, Ca2+
Elektrokjemisk potensiale for et ion kan regnes ut ved:
μ = RTln(co/ci) + zFEm
μ= potensial
R = universell gasskonstant
T= absolut ttemperatur
C = konsentrasjon på innside (i) og utside (o)
z = ionets ladning
F = Faradays konstant
Em = elektrisk potensialforskjell over membran
Ved elektrokjemisk likevekt skjer det ingen netto forflytning av ionet
Nernst likning gir likevektspotensialet til ionet (Em)
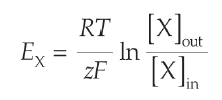
- Ionet vil diffundere mot en elektrokjemisk likevekt (gjennom ionekanaler)
- Ioner med størst permiabilitet får størst effekt på membranpotensialet
K+ som regel viktigst i kroppen (lekkasjestrøm)
Dersom likevekten var fordelt presist slik Gibbs-Donnan-modellen forutser ville vi fått et osmotisk trykk inn i cellen (pg.a. flere molekyler på innsiden). Dette får vi ikke fordi:
- Cellen tåler ikke for stor innstrømning av vann.
- Negativt ladde proteiner intracellulært balanseres av ekstracellulært Na+.
- Lekkasje av Na+ over membranen pumpes ut igjen av Na+/K+-atpase (som er viktig for reguleringen av cellevolum).
Hvordan kan celler motvirke akutte volumforandringer?
- Eksocytose
Spleise direkte med cellemembranen (totalspleising, ikke kyss og bail)
Om Na+/K+-ATPase hemmes, f.eks. av ouabain, vil cellene svelle - Akkumulering av nøytrale aminosyrer inni cellen
Produseres ved cellemetabolisme og påvirker ikke enzymfunksjon slik økte Ca2+/Na+-nivåer gjør - Regulering av vannkanaler (akvaporiner)
Foreleser: Linda Hildegard Bergersen
Ressurser
Presentasjon


