

En kuleform har minst overflate i forhold til volum. Det er derfor vannmolekyler i et fritt miljø naturlig vil orientere seg i dråper (kuler).
Amfipatiske lipider er lipider som både er hydrofobe og hydrofile (såpebobler, cellemembraner).
- Amphi (gresk): begge slag
- Pathos (gresk): følelse for noe
I en såpeboble er lipidene orientert slik at de hydrofile hodene vender mot vann og de hydrofobe mot luft.
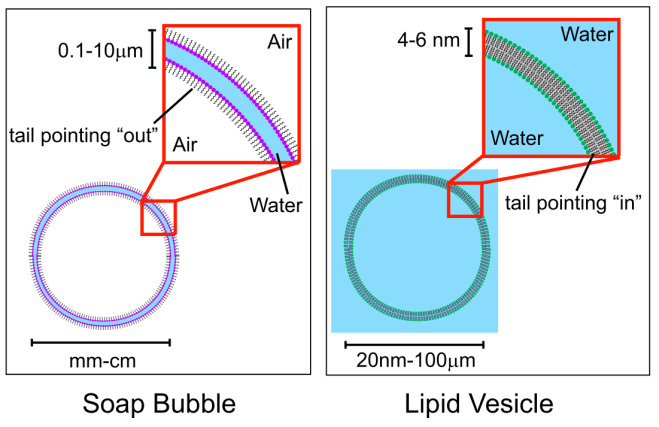
Cellemembranen er en dobbeltmembran av amfipatisk lipid.
Artige kulepunkter
- Alifatiske lipider har kjeder av karbon, vanligvis ikke forgrenede
- Lipider finnes som både mettet og umettet
Umettede lipider har dobbeltbindinger - Lipider finnes som essensielle og ikke-essensielle
Kroppen kan produsere ikke-essensielle lipider selv - Nomenklaturen er basert på IUPAC
Både systematiske og trivialnavn - C:D
C = antall karbonatomer
D = antall dobbeltbindinger - Omega (ω) – x hvor x er posisjonen til dobbeltbindingen talt fra ω-karbonatomet (siste)
F.eks. for omega-3 ligger dobbeltbindingen mellom 3. og 4. karbonatom fra omegakarbonet - Vi har cis- og transumettede fettsyrer
Orientering ved dobbeltbinding - α(alfa)-karbonet er nr. 2 og β(beta)-karbonet er nr. 3
Talt fra funksjonell gruppe
ω(omega)-karbonet er siste karbonatom
Dobbeltbindinger forekommer ofte regelmessig (f.eks. annenhver binding).
Amfipatiske lipider i vann vil selv organisere seg til en dobbeltmembran og lukke seg fordi det er en energetisk gunstig konfigurasjon. Det er ugunstig å ha hydrofobe ender vendt mot vann.
Strukturen kan være: alkohol (kolin), fosfat, glyserol, og fettsyrer (med knekk dersom det er en cis-dobbeltbinding). Ladningene på fettsyrene kan ha noe å si, men de pleier å være nøytrale.
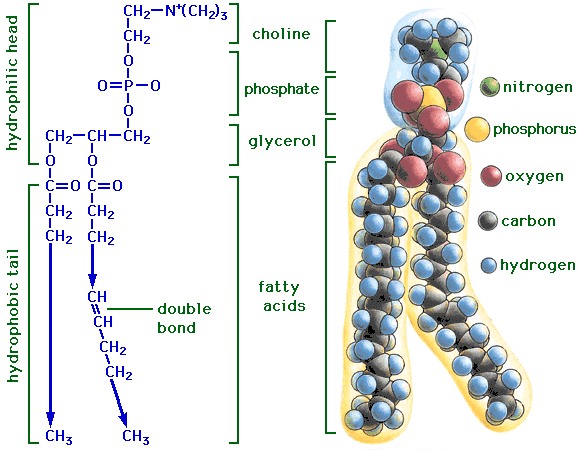
Fosfolipider er mobile. De er ikke som margarin eller oljer, men en mellomting. Fluiditeten til cellemembranen avhenger av sammensetningen (temperatur, fettsyrenes halelengde, dobbeltbindinger (flere jo mykere, lavere smeltepunkt), mengde kolesterol (lettere å flipfloppe, gjør membranen stivere pg.a. en stiv ringstruktur) usw.). Kolesterol i beskjedne konsentrasjoner reduserer fluiditeten, men øker i høy (hos røde blodceller består membranen av nesten ~1/4 kolesterol). Membranen blir stivere jo lengre unna sentrum du kommer (men hvor er sentrum?). Tiltrekningen mellom lipidhalene øker med lengden da de induserte londonkreftene blir sterkere. Dipolkraften avtar med d^6 (d = distance). (Enkelte) Bakterier kan justere fettsyresammensetningen i membranene deres med et ytre miljø (bedre tilpasning).
Cellemembranen har god lateral (sidelengs) mobilitet. Lipidene kan flyte sidelengs som en “todimensjonal væske” En “flip” hos lipidene kan skje (at de bytter plass), men veldig sjeldent da det polare hodet da må passere gjennom det hydrofobe indre partiet.
På membranen finner vi (blant annet) mange vannporer (akvaporiner). Vi trenger dem fordi osmose som eneste transportmekanisme hadde vært for treg. Den nøyaktige sammensetningen av lipidmembranen er ukjent (f.eks. hos gliaceller). Har vi mye kolesterol blir hjernen mindre permiabel for vann.
De to lagene i dobbeltmembranen har ulik sammensetning (pg.a. enzymer, flippaser). Membraner dannes i endoplasmatisk retikulum. Glykolipider finnes bare i det ytre laget og er viktig for å beskytte cellen mot uvennlige ytre omgivelser (lav pH, nedbrytningsenzymer usw.). Adhesjon mellom celler er også avhengig av glykolipider. I det innerste laget (vendt mot cytosol) finner vi typisk negativt ladde ioner. De fleste glykolipider er basert på sfingosin (samme som sfingomyelin). Gangliosid er en overflatereseptor for bakterietoksinet som forårsaker diare ved kolera. Nedbryting av fosfolipider kan være viktig for å overføre signaler fra det ekstracellulære miljøet til cellens lumen (intracellulært).
Foreleser: Erlend Arnulf Nagelhus
Ressurser
Presentasjon


{kind=link}